DNA – Livets oppskrift
DNA, eller deoksyribonukleinsyre, er selve bæreren av genetisk informasjon i nesten alle levende organismer. Det er dette molekylet som inneholder «oppskriften» på hvordan celler skal bygges, fungere og formere seg. Alt fra øyefarge til hvordan leveren skal fungere, ligger kodet i DNA – et molekyl så lite at det ikke kan sees med vanlig mikroskop, men med en informasjonsmengde som overgår det meste vi kjenner til.
Oppbygning av DNA – molekylets struktur
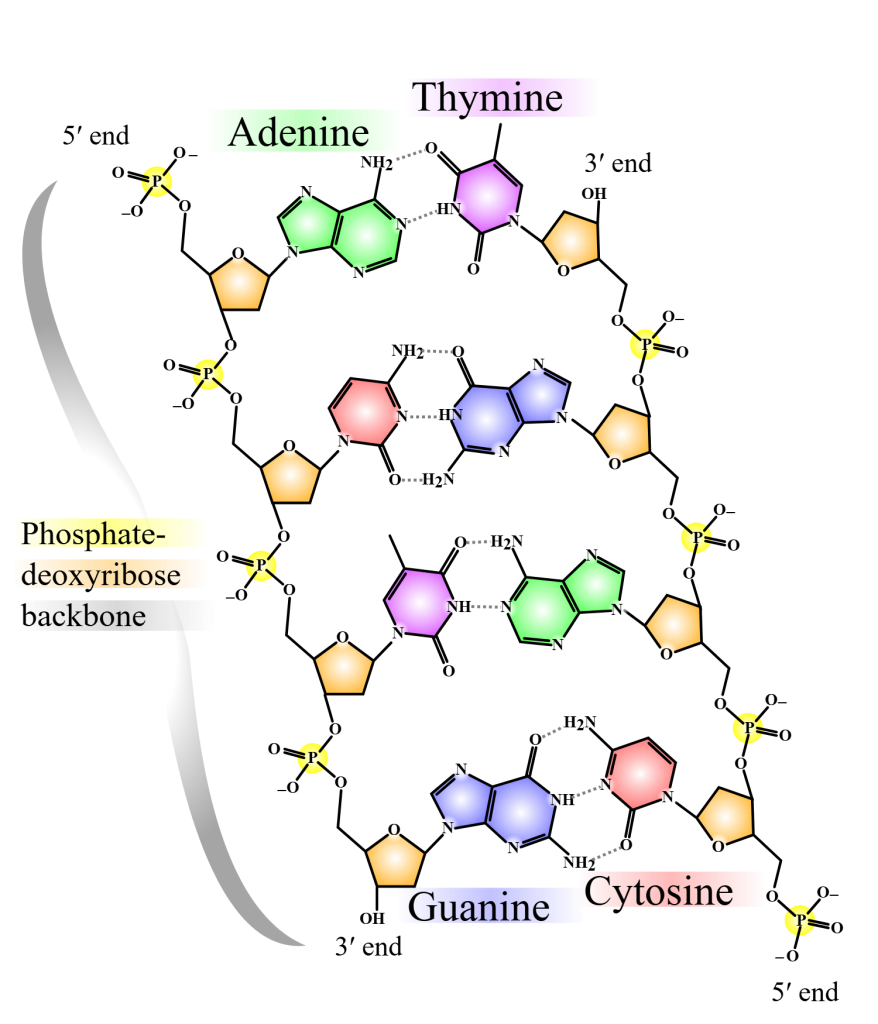
DNA er bygget opp som en dobbelheliks – to lange kjeder som snor seg rundt hverandre, litt som en vridd stige. Hver «side» av stigen er laget av vekselvis sukker og fosfat, mens «trinnene» utgjøres av basepar som holder de to kjedene sammen.
Byggesteinene: Nukleotidene
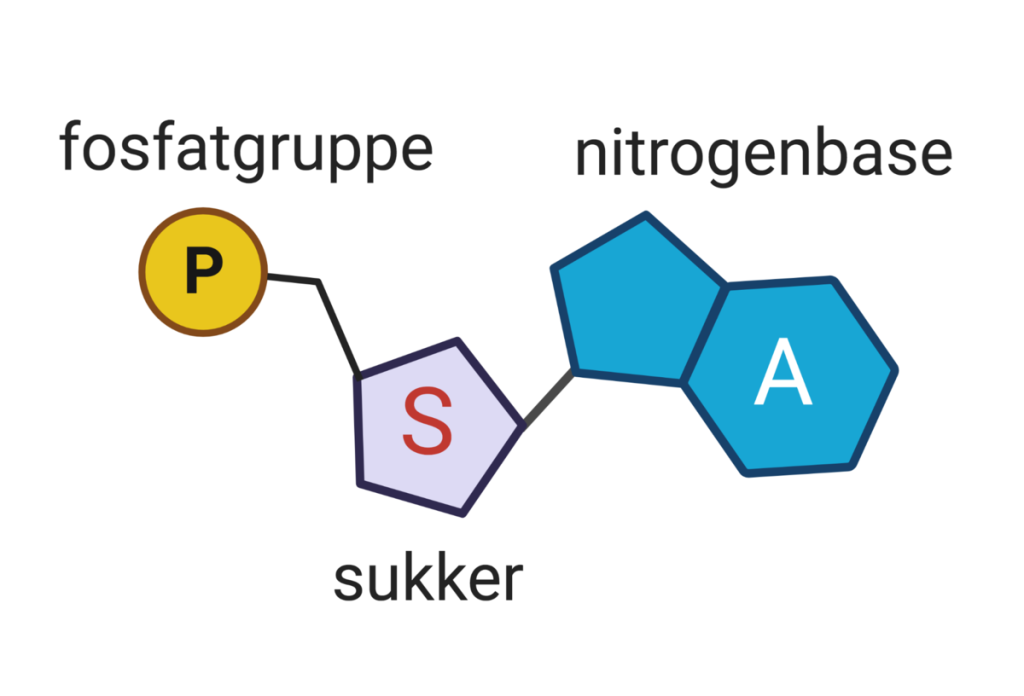
Grunnenheten i DNA er nukleotidet. Hvert nukleotid består av tre deler:
- Et sukkermolekyl – deoksyribose
- En fosfatgruppe
- En nitrogenbase
Det finnes fire ulike nitrogenbaser i DNA:
- Adenin (A)
- Tymin (T)
- Guanin (G)
- Cytosin (C)
Disse basene parer seg med hverandre på en bestemt måte, et prinsipp kalt komplementær baseparring:
- Adenin (A) binder seg alltid til Tymin (T)
- Guanin (G) binder seg alltid til Cytosin (C)
Bindingene mellom baseparene holdes sammen av hydrogenbindinger. A–T har to slike bindinger, mens G–C har tre. Det gjør at G–C-parene er litt sterkere og bidrar til økt stabilitet der de finnes i stor konsentrasjon – for eksempel i gener som trenger høyere termisk stabilitet.
Retningen i DNA – forståelsen av 5’ og 3’
Et DNA-molekyl er ikke bare en tilfeldig snodd tråd. Det er et strukturert, retningsbestemt system der hvert molekyl har en begynnelse og en slutt – en retning. Dette kan sammenlignes med å lese en bok: Det har betydning hvor du starter, og i hvilken rekkefølge du leser setningene. På samme måte må DNA leses eller kopieres i en bestemt retning – og denne retningen bestemmes av strukturen i DNA-trådene.
Byggesteinen: nukleotidet og sukkerets orientering
Hver DNA-tråd består av mange små byggesteiner som kalles nukleotider.
Hvert nukleotid består av tre deler: et sukkermolekyl (deoksyribose), en fosfatgruppe og en nitrogenbase (A, T, G eller C).
Det er strukturen på sukkeret som bestemmer hvordan nukleotidene lenkes sammen, og som gir DNA-tråden dens retning.
Sukkeret i hvert nukleotid har fem karbonatomer.
Disse nummereres fra 1’ til 5’ (uttales «én merket» til «fem merket»).
Disse tallene brukes som referanse for å beskrive hvilke deler av molekylet som deltar i bindingene mellom nukleotidene:
- 5’-enden (5-prime) er den enden av tråden hvor fosfatgruppen er bundet til karbon nummer 5 i sukkeret.
- 3’-enden (3-prime) er den enden hvor en hydroksylgruppe (–OH) er festet til karbon nummer 3.
Når nye nukleotider kobles til en voksende DNA-tråd, skjer det alltid ved at fosfatgruppen på det nye nukleotidet kobles til hydroksylgruppen på 3’-karbonet på det forrige. Dermed vokser tråden i én bestemt retning: fra 5’ til 3’.
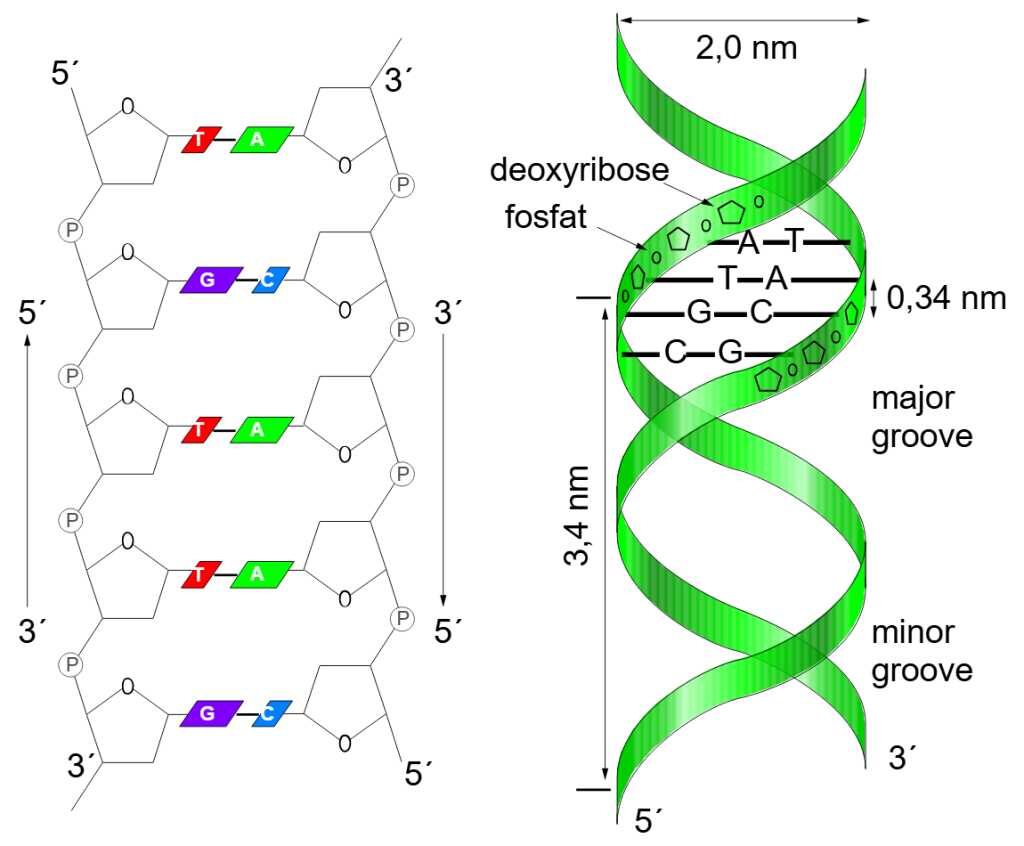
Antiparallelle tråder – to tråder i motsatt retning
Et DNA-molekyl består av to slike tråder som snor seg rundt hverandre i en dobbelheliks. Men de to trådene går i motsatt retning. Den ene går fra 5’ til 3’, og den andre fra 3’ til 5’. Dette kalles at DNA-trådene er antiparallelle.
Dette arrangementet er ikke tilfeldig. Det er helt nødvendig for at baseparingen skal fungere. A binder til T, og G til C, gjennom hydrogenbindinger – og disse bindingene krever at molekylene ligger i speilvendt orientering.
Du kan tenke på det som en glidelås: Tennene på hver side må stå speilvendt for å kunne kobles sammen. Det samme gjelder DNA.
Enzymenes retning: hvorfor 5’ til 3’ betyr noe
Mange av prosessene som skjer i cellen – som kopiering av DNA før celledeling, eller avlesing av DNA når det skal lages proteiner – er avhengige av spesifikke enzymer. Disse enzymene, som DNA-polymerase og RNA-polymerase, kan bare arbeide i 5’ til 3’-retningen.
Dette har store konsekvenser:
- Når DNA skal kopieres (replikasjon), brukes den ene tråden som en mal i 5’ til 3’-retning, og den andre i 3’ til 5’-retning. Men siden enzymene bare kan lage ny tråd i 5’ til 3’-retning, oppstår en asymmetri.
- Den ene tråden – kalt leading strand – kopieres jevnt og kontinuerlig.
- Den andre tråden – kalt lagging strand – må kopieres stykkevis i små biter (kalt Okazaki-fragmenter), som senere limes sammen.
Et stabilt, men dynamisk molekyl
Selv om DNA er ekstremt stabilt – det kan bevares i tusenvis av år under riktige forhold – er det også et dynamisk molekyl. Det åpnes og lukkes hele tiden i cellen. Når et gen skal leses, «lynes» trådene opp i det aktuelle området, slik at informasjonen i basekodene kan oversettes til RNA og deretter til proteiner.
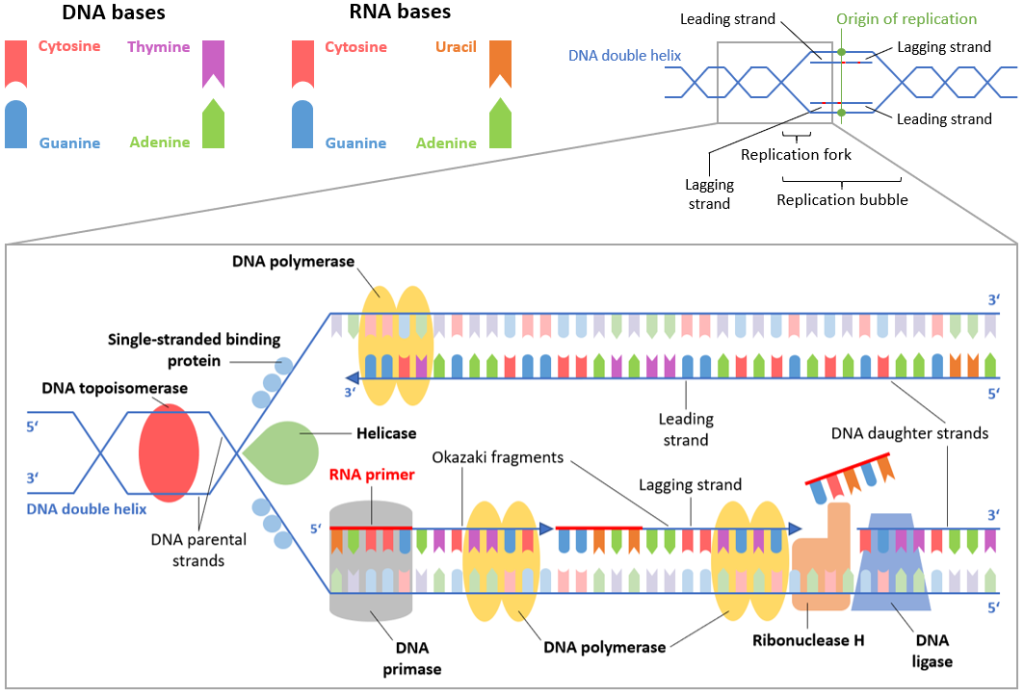
Replikasjonens start: Hvordan DNA kopieres
DNA-replikasjonen er en presis og kontrollert prosess der cellen lager en nøyaktig kopi av sitt eget arvestoff før celledeling. Denne prosessen starter alltid på bestemte steder på DNA-tråden, kalt replikasjonsorigins – eller på norsk, replikasjonsoriginer. Disse er korte DNA-sekvenser som fungerer som startpunkter der replikasjonsmaskineriet kan feste seg og begynne å arbeide.
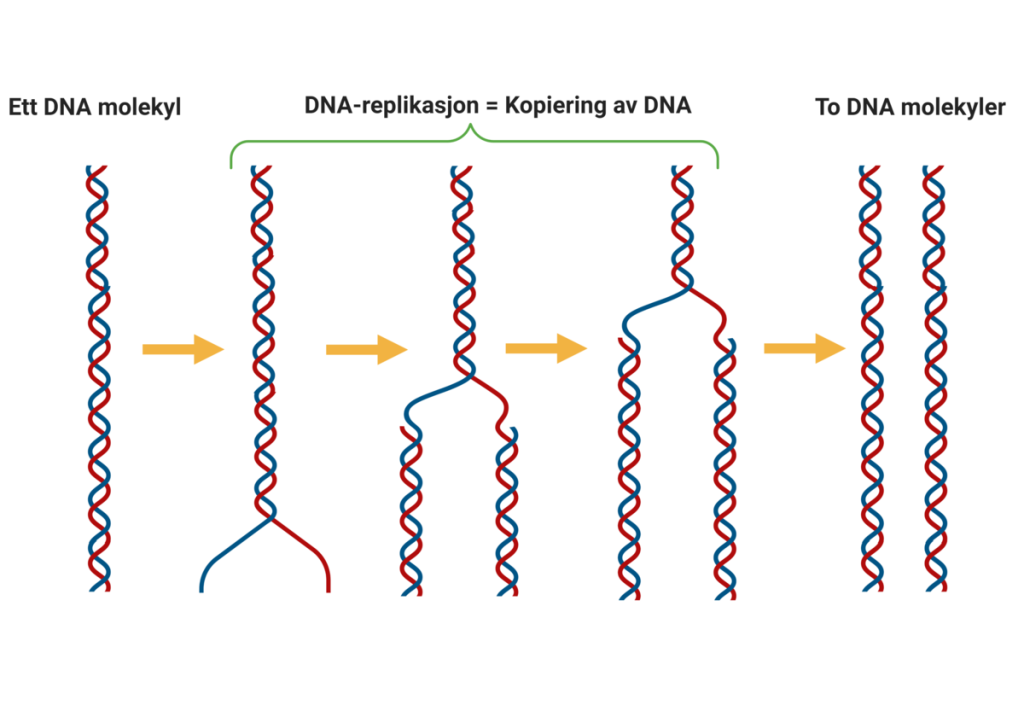
Åpning av DNA-dobbelheliksen
Når replikasjonen skal i gang, trer enzymet helikase i aksjon. Helikase fungerer som en «molekylær glidelås» som åpner opp dobbelheliksen ved å bryte hydrogenbindingene mellom baseparene – litt som å dra fra hverandre en glidelås.
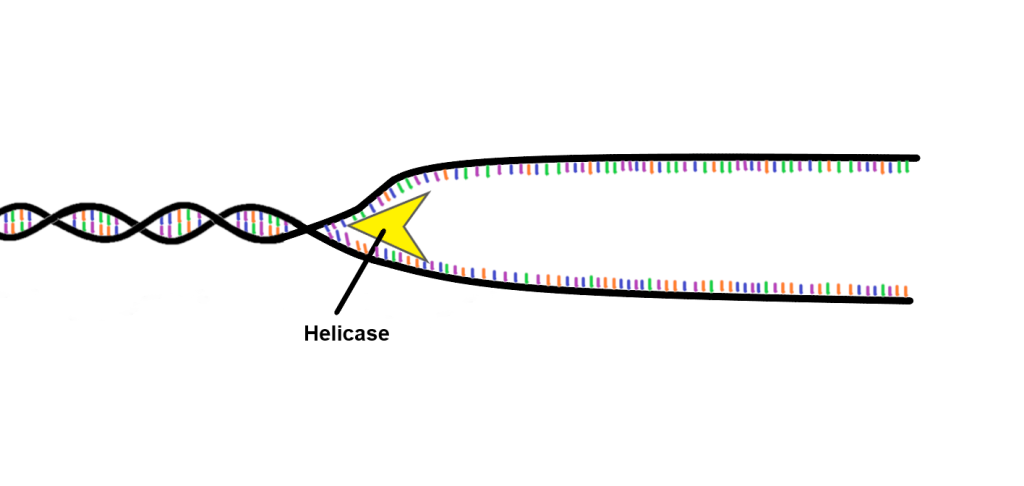
Dette skaper en Y-formet struktur vi kaller replikasjonsgaffelen. I denne gaffelen åpnes DNA kontinuerlig i begge retninger fra origo, og vi får en bidireksjonell replikasjon.
For å hindre at de enkeltrådede DNA-strengene umiddelbart binder seg sammen igjen, binder egne proteiner – såkalte single-strand binding proteins (SSB) – seg til trådene.
Disse holder trådene adskilt og beskytter dem mot nedbrytning mens replikasjonen pågår.
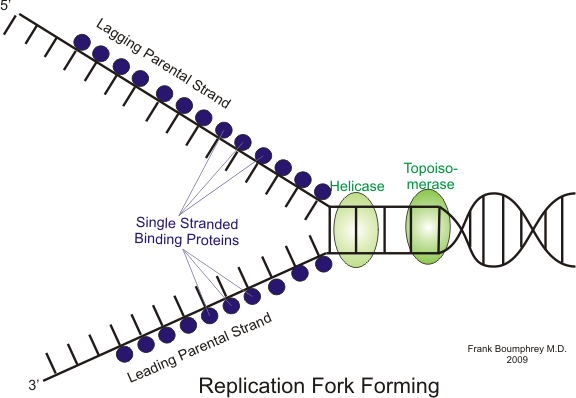
Samtidig oppstår det et fysisk problem: Når helikase åpner dobbelheliksen, skapes det spenninger og overvridning lenger oppe på DNA-tråden.
Dette kan føre til at DNA kveiler seg for stramt, en tilstand kjent som supercoiling. For å løse dette, kommer topoisomeraser inn og kutter DNA-et midlertidig, vrir det og limer det sammen igjen – som når man løser en floke ved å løsne og justere trådene. Topoisomerase I kutter én tråd om gangen, mens Topoisomerase II kan kutte begge trådene samtidig og takle mer komplekse floker.
Bygging av ny DNA-tråd: Enzymenes samspill
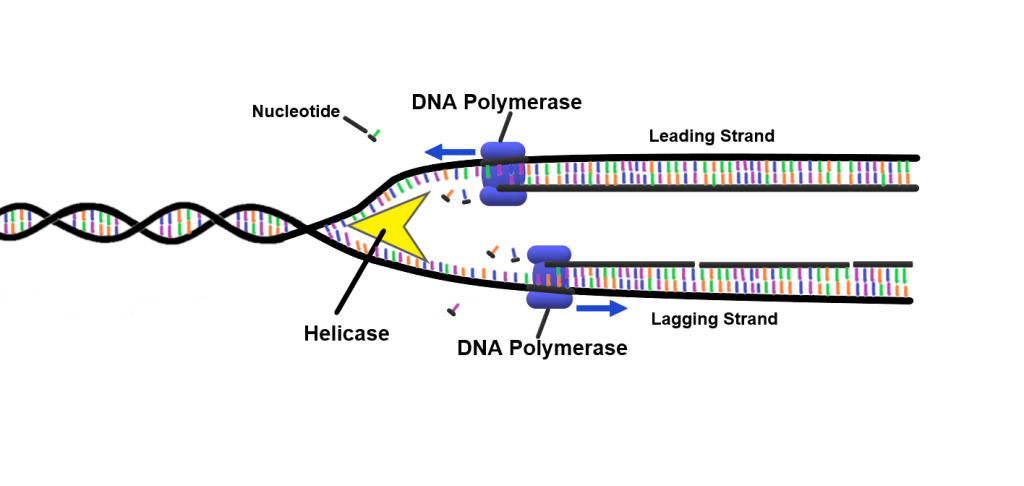
DNA-polymerase – enzymet som faktisk bygger nye DNA-tråder – har en viktig begrensning: den kan ikke starte på egen hånd. Den trenger en startplattform.
Derfor kalles enzymet primase inn.
Primase lager korte RNA-biter, såkalte primere, som gir polymerasen et utgangspunkt.
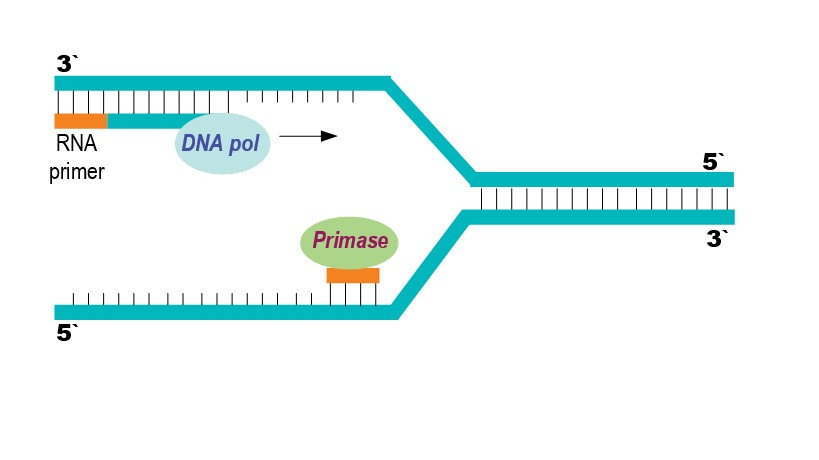
Når RNA-primeren er lagt, tar DNA-polymerase alfa over og forlenger primeren med en kort DNA-sekvens.
Dette fungerer som en overgang, en slags «startmotor» før de mer effektive polymerasene overtar.
To spesialiserte DNA-polymeraser gjør deretter hovedjobben: DNA-polymerase epsilon jobber på den ledende tråden og kan følge helikasen i én lang, sammenhengende bevegelse.
På den andre siden av replikasjonsgaffelen, hvor DNA går i motsatt retning, må DNA-polymerase delta jobbe mer stykkevis.
Her dannes korte DNA-biter kjent som Okazaki-fragmenter, som etter hvert blir limt sammen til én lang tråd.
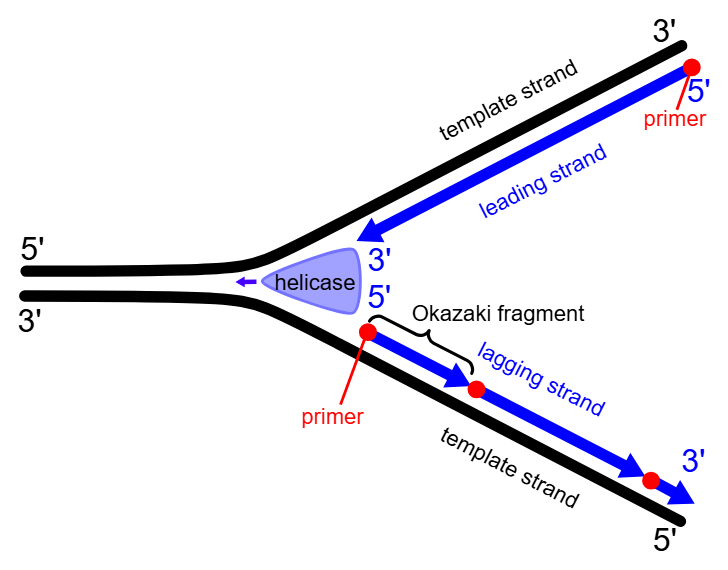
Denne oppdelingen skyldes DNA-molekylets antiparallelle natur: de to trådene løper i motsatt retning (fra 5’ til 3’ og fra 3’ til 5’).
Fordi DNA-polymerase bare kan jobbe i 5’ til 3’-retning, blir den ene tråden replikert kontinuerlig (den ledende tråden), mens den andre må bygges i små, diskontinuerlige biter (den etterslepende tråden).
Test deg selv
Koble sammen og rydde opp
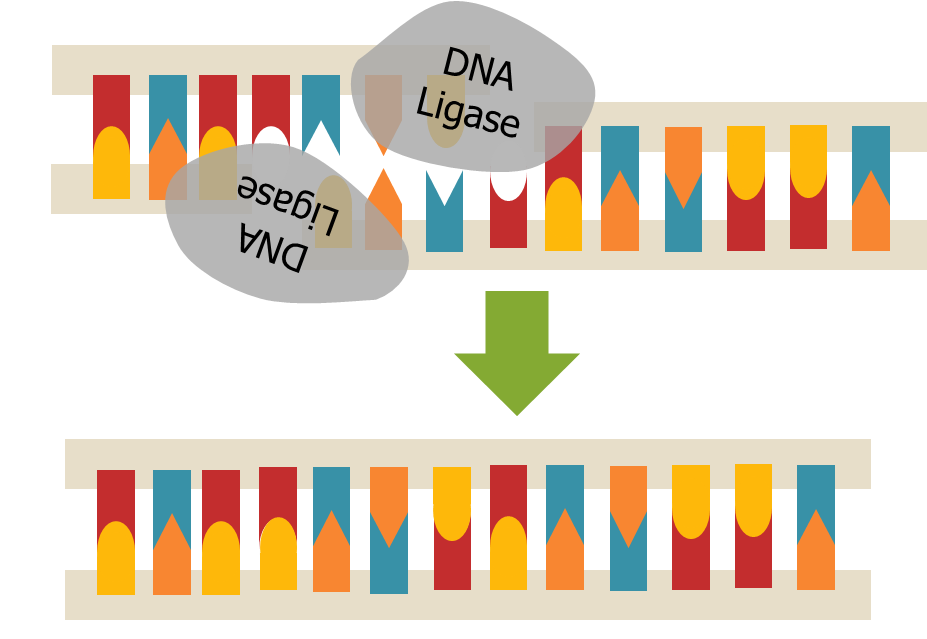
Når alle Okazaki-fragmentene er ferdigsyntetisert, må de kobles sammen til én hel DNA-tråd.
Dette gjøres av enzymet DNA-ligase, som fungerer som cellens egen «limstift».
Den lukker hullene mellom fragmentene og gjør den nye DNA-tråden sammenhengende.
Sikkerhet og presisjon: Hvordan unngås feil?
For å sikre at DNA kopieres korrekt, bygger DNA-polymerase på prinsippet om komplementær baseparing: A parer alltid med T, og G med C. Men i tillegg til dette har enzymet innebygd korrekturlesing – såkalt proofreading.
Hvis et feil nukleotid settes inn, kan polymerasen oppdage dette, fjerne feilen og sette inn riktig base før den går videre.
Dette gjør DNA-replikasjonen svært nøyaktig – faktisk skjer det bare én feil per milliard baser!
Et viktig tillegg til nøyaktigheten er det som kalles «induced fit» – når polymerasen bare endrer form og godtar nukleotidet dersom det passer perfekt inn i den aktive lommen.
Det gir en ekstra seleksjonsbarriere mot feil.
Viktige polymeraser å huske:
| Polymerase | Funksjon | Hvor i cella? |
|---|---|---|
| Alfa (α) | Initierer DNA-replikasjon ved å forlenge RNA-primere med en kort DNA-sekvens | Cellekjernen |
| Beta (β) | Hovedsakelig involvert i DNA-reparasjon | Cellekjernen |
| Delta (δ) | Hovedansvarlig for syntese av den etterslepende tråden og Okazaki-fragmenter | Cellekjernen |
| Epsilon (ε) | Primært ansvarlig for syntese av den ledende tråden | Cellekjernen |
| Gamma (γ) | Involvert i replikasjon av mitokondrielt DNA | Mitokindriene |
Eksamensoppgave
DNA-pakking – hvordan 2 meter arvemateriale får plass i cellekjernen
I hver menneskecelle finnes det omtrent 2 meter med DNA. Likevel ligger dette enorme molekylet kompakt lagret i en cellekjerne som bare måler rundt 6 mikrometer i diameter. For å gjøre dette mulig, er DNA organisert gjennom flere nivåer av pakking – en prosess som både beskytter arvematerialet og regulerer hvilke gener som er tilgjengelige for transkripsjon.
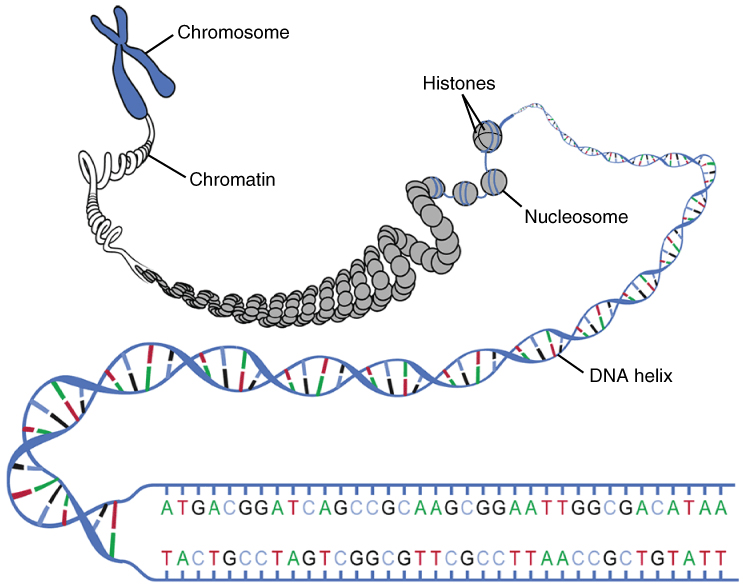
1. Den nakne dobbelheliksen – utgangspunktet for pakking
DNA starter som en tynn tråd – den velkjente dobbelheliksen – som har en diameter på omtrent 2 nanometer. Denne strukturen er stabil og lang, men ikke egnet for lagring i sin frie form. For å kunne få plass og samtidig holdes organisert og funksjonell, må den pakkes inn i et mer kompakt og strukturert format.
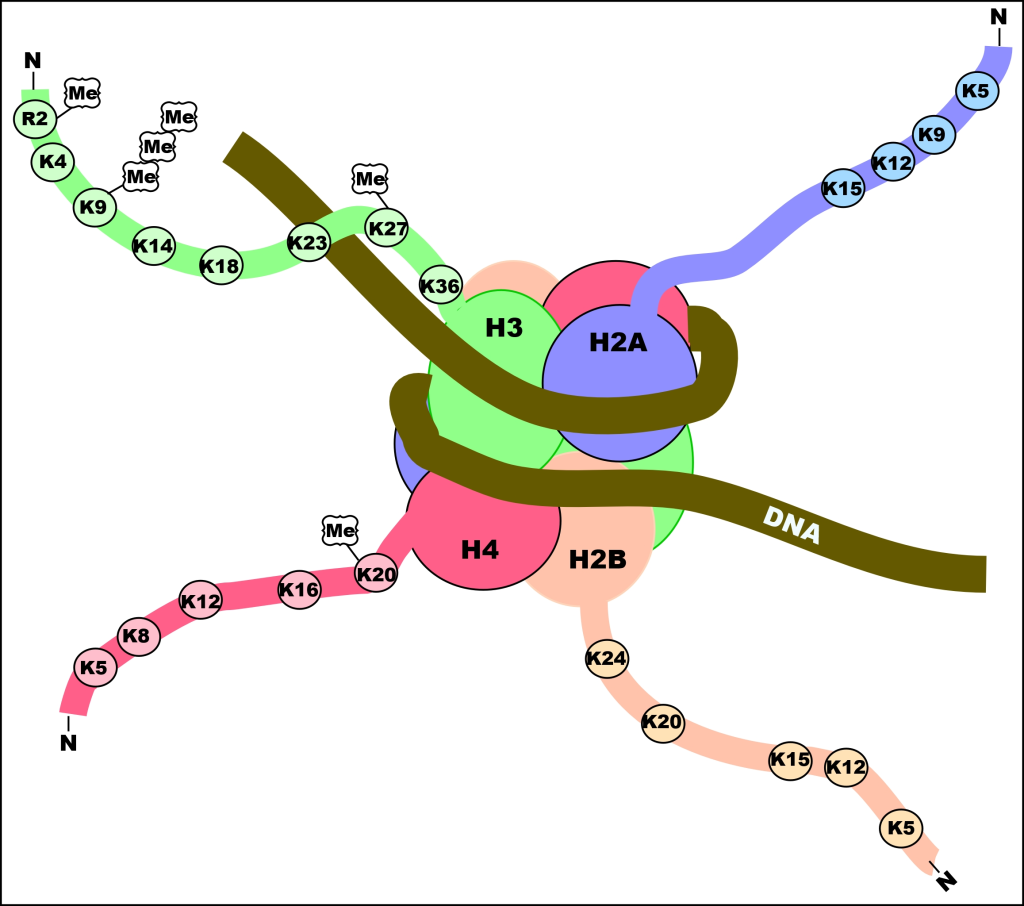
2. Nukleosomer – DNA vikles rundt histoner
Det første nivået av pakking skjer ved hjelp av proteiner kalt histoner. Disse danner en slags «spole» som DNA kan vikles rundt. Hver histon består av åtte proteinmolekyler (to av hver: H2A, H2B, H3 og H4), og rundt disse vikler DNA seg omtrent 1,65 ganger – tilsvarende 147 basepar.
Hver slik struktur kalles et nukleosom, og mellom hvert nukleosom ligger en liten DNA-sekvens kalt linker-DNA. Dette gir en struktur som ofte omtales som «perler på en snor».
På dette stadiet har DNA en tykkelse på omtrent 11 nanometer. Det er fortsatt delvis tilgjengelig for transkripsjon, og dette nivået dominerer i celler som er i aktiv bruk av sine gener.
3. 30-nanometer-fiberen – tettere organisering
Nukleosomstrukturen foldes videre og danner en tykkere fiber med en diameter på omtrent 30 nanometer. Her spiller et ekstra histon, H1, en viktig rolle. Det binder seg til linker-DNA og hjelper med å trekke nukleosomene tettere sammen.
Denne tettere strukturen kalles ofte solenoide eller zigzag-fiber, avhengig av hvordan nukleosomene organiseres. På dette nivået begynner DNA å bli mindre tilgjengelig for transkripsjon, og det markerer overgangen til et mer kompakt, men fortsatt dynamisk, kromatin.
4. Kromatin – funksjonell DNA-pakking
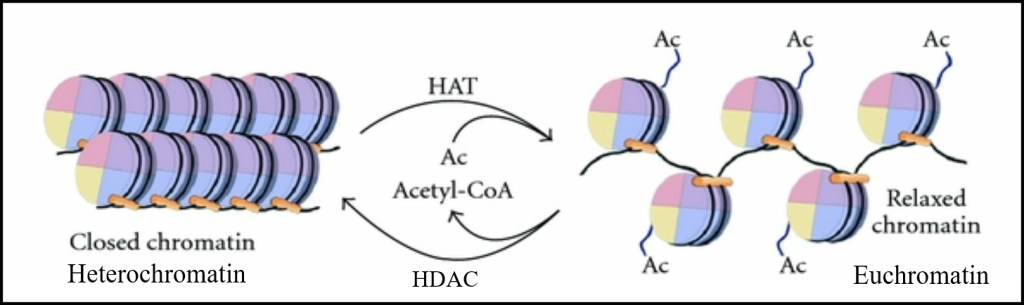
Det vi ser i cellekjernen utenfor celledeling kalles kromatin. Kromatin finnes i to hovedformer:
- Eukromatin: Løst pakket, transkripsjonelt aktivt DNA – her kan genene leses.
- Heterokromatin: Tett pakket, transkripsjonelt inaktivt DNA – her er genene stort sett «avskrudd».
Denne differensieringen gjør at cellen kan styre hvilke deler av genomet som skal brukes, og hvilke som holdes inaktive. Det er avgjørende for celledifferensiering og genregulering.
5. Kromatinløkker og høyere ordens struktur
Kromatinfiberen på 30 nanometer danner videre strukturer ved å foldes i løkker, som festes til et slags protein-skjelett inne i cellekjernen. Disse kromatinløkker kan samles i domenestrukturer, og på denne måten organiseres genene i funksjonelle enheter.
Når cellen ikke deler seg, flyter dette organiserte kromatinet fritt rundt i kjernen som et løst, men strukturert nettverk. Men når celledeling starter, skjer det en dramatisk reorganisering.
6. Kromosomer – den mest kondenserte formen
Før celledeling må DNA pakkes maksimalt for at det skal kunne fordeles korrekt til dattercellene. Kromatinfibrene kondenseres da ytterligere til kromosomer, som er synlige i lysmikroskop som X-formede strukturer under metafasen av mitosen.
Hvert kromosom består da av to identiske søsterkromatider, som holdes sammen i et område kalt sentromeren. DNA er nå ikke lenger tilgjengelig for transkripsjon – hele genomet er midlertidig «stengt», for å fokusere på kopiering og fordeling.
📌 Kort oppsummert: Nivåer av DNA-pakking
| Nivå | Struktur | Diameter |
|---|---|---|
| Dobbelheliks | Uviklet DNA | 2 nm |
| Nukleosom | DNA viklet rundt histoner | 11 nm |
| 30-nm fiber | Tettere fiberstruktur med H1-histon | 30 nm |
| Kromatin | Samlet struktur i kjernen (eukromatin/hetero) | Varierende |
| Kromosom | Maksimal pakking under celledeling | Synlig nivå |
📚 Anki-kort
Obs, tomt! Kommer etterhvert <3
📝 Eksamensoppgaver
Obs, tomt! Kommer etterhvert <3
👨⚕️ Klinisk case
Obs, tomt! Kommer etterhvert <3
❓ Test deg selv
Obs, tomt! Kommer etterhvert <3
Legg igjen en kommentar